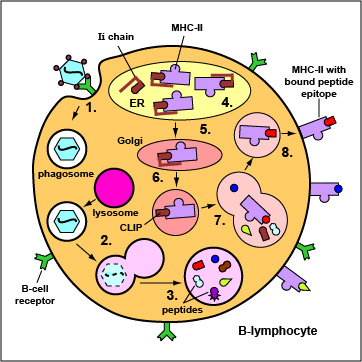
Exogenous antigens are those from outside cells of the body. Examples include bacteria, free viruses, yeasts, protozoa, and toxins. These exogenous antigens bind to B-cell receptors and enter B-lymphocytes through endocytosis. After lysosomes fuse with the phagosome, protein antigens are degraded by proteases into a series of peptides. These peptides eventually bind to grooves in MHC-II molecules and are transported to the surface of the B-lymphocyte. Effector T4-lymphocytes are then able to recognize peptide/MHC-II complexes by means of their T-cell receptors (TCRs) and CD4 molecules.
1. Epitopes of exogenous antigens, such as viruses, bind to a complementary shaped B-cell receptor on a B-lymphocyte. The antigen is engulfed and placed in a phagosome.
2. Lysosomes fuse with the phagosome forming an phagolysosome.
3. Protein antigens are degraded into a series of peptides.
4. MHC-II molecules are synthesized in the endoplasmic reticulum and transported to the Golgi complex. Once assembled, within the endoplasmic reticulum, a protein called the invarient chain (Ii) attaches to the the peptide-binding groove of the MHC-II molecules and in this way prevents peptides designated for binding to MHC-I molecules within the ER from attaching to the MHC-II.
5. As the MHC-II molecules with bound Ii chain are transported to the Golgi complex, the Ii is cleaved, leaving a short peptide called CLIP in the groove of the MHC molecule.
6&7. The vesicles containing the MHC-II molecules fuse with the peptide-containing phagolysosomes. The CLIP peptide is removed from the MHC=II molecules and the peptide epitopes are now free to bind to the grooves of the MHC-II molecules.
8. The MHC-II molecules with bound peptides are transported to the cytoplasmic membrane of the B-lymphocyte where they become anchored. Here, the peptide and MHC-II complexes can be recognized by effector T4-lymphocytes by way of TCRs and CD4 molecules having a complementary shape.
Last updated: August, 2019
Please send comments and inquiries to Dr.
Gary Kaiser
